Pheasant Phylogenetics: an Overview of my Findings
Krissy Bush
Department of Biological Sciences, University of Alberta
kbush@aviangenetics.com
Pheasants are one of the most endangered groups of birds in the world. A total of 27 species appear on the most recent Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES) list, with 17 of these classified as CITIES I (Appendix I species are either rare or endangered). In the past 150 years, several pheasant species and subspecies have virtually disappeared with few birds left in the wild and limited breeding stock in captivity. Accurate evolutionary and taxonomic information is therefore essential for developing appropriate CITES classifications and for the management of threatened species.
What exactly is a pheasant? To date, the boundaries that define pheasants have not been satisfactorily determined. Pheasants are generally thought to be comprised of three of the subfamilies present in Phasianidae: Tragopaninae (Tragopans), Argusianinae (Argus Pheasants), and Phasianinae (Pheasants). Tragopaninae is a single genus that consists of the five Tragopan species and Argusianinae is made up of the three genera, Polyplectron (Peacock Pheasants), Rheinartia (Crested Argus), and Argusianus (Great Argus). Phasianinae is much more diverse and encompasses 10 different genera: Ithaginis (Blood Pheasants), Pucrasia (Koklass), Lophophorus (Monal Pheasants), Gallus (Junglefowl), Lophura (Gallo Pheasants), Crossoptilon (Eared Pheasants), Catreus (Cheer), Syrmaticus (Long-Tailed Pheasants), Phasianus (True Pheasants), and Chrysolophus (Ruffed Pheasants). This is however a mainly arbitrary classification used to group together genera that are similar in body form and behavior.
There have been several recent attempts to reconstruct phylogenetic relationships of the Phasianidae, but a comprehensive study including all three subfamilies (Tragopaninae, Argusianinae, and Phasianinae) has not been attempted. The present study was inspired by Kimball et al. (1999) who examined the molecular phylogeny of pheasants and partridges. Their main goal was to determine whether these subfamilies are monophyletic (a single evolutionary group where all of the members evolved from a single common ancestor). They sequenced the complete cytochrome b gene of various species from multiple genera of galliformes, but sampled only one species from each genus of pheasants commonly found in captivity. We decided to sequence the cytochrome b region of all pheasant species readily available in Canada in order to try to further resolve their phylogenetic relationships.
The primary goal of this study was to ascertain the evolutionary relationships among the different species of pheasants and to determine whether the current taxonomic classification is correct or needs to be revised. Other secondary considerations were to determine if the pheasant group is monophyletic (do all of the members of this group share a common ancestor and does that ancestor only give rise to members of the named taxon), which has been disputed in several previous studies. A final goal was to determine if the Grey Peacock Pheasant is a pheasant or a peafowl. It shares characteristics with both groups, but is traditionally classified as a pheasant because of its geographic range and morphological features.
Results
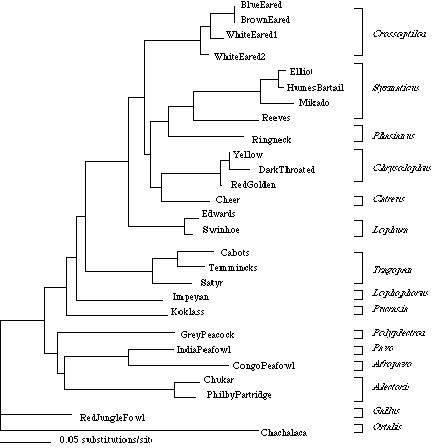
Most groups stay constant throughout all forms of phylogenetic analysis (Crossoptilon, Syrmaticus/Phasianus, Chrysolophus, Lophura, Tragopan, Pavo/Afropavo, and Alectoris), but four species, the Cheer, Red Junglefowl, Koklass and Grey Peacock, are highly variable in their placement.
Placement of the Grey Peacock Pheasant within Phasianidae
Traditional studies have placed the Grey Peacock with either the pheasants or the peafowl depending on the classification approach. The Peacock pheasants (Genus Polyplectron) have always been classified as a pheasant because of their characteristics, but received the name peacock because of their eyed feathers and display. The assignment of the Grey Peacock as a pheasant seems reasonable because of their behavior and appearance, but recent phylogenetic studies have placed them with the peafowl.
In one of our types of phylogenetic analysis (maximum likelihood), the Grey Peacock pairs with the Peafowl-Partridge grouping (Figure 1). In the other type of phylogenetic analysis (maximum parsimony), it groups with the two Old-World Partridges. Since the Grey Peacock does not consistently pair with one group, it can not be ruled out as being a pheasant with any certainty, but it is becoming increasingly unlikely with the mounting evidence (Kimball et al. 1999 and this study). It paired with both the partridges and the peafowl in this study so it is likely that it may be a member of either group. Johnsgard hypothesized that many or all of the typical pheasant lineages evolved from a partridge-like ancestor, but there is no data to support that. However, this theory may explain why a pheasant-like bird is grouping basally with the partridges. To determine whether the Grey Peacock belongs with the partridges or peafowl, more data is necessary. More species of Peacock Pheasants must be sequenced, along with more species of peafowl and partridge.
Placement of Catreus wallichi (Cheer) within the Pheasants
Cheer are a very unique pheasant therefore using morphological data to try resolve their placement is not very productive. They share characteristics with every group, but do not fall into any clear category historically because of their odd appearance and mix of morphological and behavioral traits. Both sexes are relatively similar with regards to appearance, which is unusual in the pheasants outside the Eared, and they are both a very subdued sand colour, which is uncharacteristic for male pheasants. Perhaps due to their unusual characteristics, it is not surprising that Cheer was highly mobile when different methods of analysis were used. Maximum likelihood resulted in the Cheer pairing with the Ruffed group (Figure 1) while maximum parsimony causes the Cheer to pair with the Reeves and Ringneck. In a previous studies Cheer were placed with the Red Goldens and the White Eared Pheasants or with the Ruffed clade. Again, more species and more genes are needed to resolve the phylogenetic relationships of the pheasants.
The Placement of the Genus Pucrasia (Koklass) within Phasianidae
Pucrasia is a highly distinctive genus whose placement remains unresolved. Bootstrapping (a third tool in phylogenetic analysis) positions Koklass basal to the monophyletic pheasant grouping. Maximum likelihood places them either as the most primitive pheasant or as the sister (most closely related) to the whole pheasant lineage (Figure 1). The most interesting placement occurs when maximum parsimony was attempted with altered character weights where Koklass becomes nested in the the non-pheasant lineages as the sister species to all pheasants, peafowl, and partridges. These different placements within the tree raise questions as to whether the Koklass is actually a pheasant and if it is, are pheasants monophyletic. Due to the uncertainty of classifying either the Grey Peacock or the Junglefowl as pheasants, it makes it difficult to determine whether the Koklass is a pheasant. With the present data, it is exceedingly unlikely that the Koklass should be included with the pheasants and therefore pheasants are likely monophyletic. More mitochondrial data, as well as nuclear markers, should be sequenced for all of the Koklass species along with other pheasant species, to try determine the phylogenetic relationship of this genus.
The Placement of the Genus Gallus within Phasianidae
Gallus is traditionally included with the pheasants in the subfamily Phasianinae however in all forms of analysis, they appear to be ancestral to the pheasant lineage. When boot strapping was performed, the Red Junglefowl was located between the monophyletic pheasant group and the partridge-peafowl group along with the Grey Peacock and Koklass whose placement also remains unresolved. In both the maximum likelihood and maximum parsimony trees (Figure 1), The Red Junglefowl appears to be the sister group to the outgroup (the Chachalaca). It is therefore likely that the genus Gallus is not a member of the pheasants, but rather shared a basal ancestor with them. To try to further resolve their relationship, all three genera of Gallus need to be included in any future analyses.
The Great Sequence Divergence of the Black Throated Golden
The Black Throated Golden is thought to be a colour mutation of the Red Golden Pheasant first developed in 1865. Maximum likelihood analysis shows the Black Throated Golden diverging from the Red Golden along with the Yellow Golden (Figure 1). Both boot strapping and maximum parsimony have the Black Throated Golden diverging from a common ancestor shared with the Red Golden making them sister taxa. The likelihood tree is consistent with the classical idea of it being a colour mutation, however the degree of sequence divergence between the Black Throated and Red Goldens (26 base pair changes) is very unlikely to have developed in only 137 years. There have been two schools of thought on the Golden colour varieties. The first and most popular is that they are all natural colour mutations of the Red Golden, but there is no scientific literature detailing specific pedigrees to support this. The actual inheritance patterns outlined for the Black Throated Golden also do not conform to any known accepted inheritance patterns and the lineage does not consistently produce birds with Black Throated characteristics. The second, and more probable theory, is that the Black Throated Golden is the result of selective cross breeding with the Amherst Pheasant. This theory is based on comparing feather patterns and colours of the Amherst with the Black Throated which are not present on the Red Golden. Since we were unable to sequence any Amherst sample, we can not support or disprove either hypothesis pertaining to the origination of the Black Throated lineage. The full sequence of the Amherst and the Salmon Golden is needed to determine the placement of the Black Throated Golden. The Salmon Golden is supposedly the result breeding a Black Throated Golden to a Yellow Golden. Further work to try resolve this question is being presently conducted.
Conclusions
The phylogeny of the three subfamilies (Tragopaninae, Argusianinae, and Phasianinae) of pheasants remains incompletely resolved. The placement of Cheer within the group is uncertain and it is unclear whether the Grey Peacock, Koklass, or Red Junglefowl are pheasants. Since their respective positions were unstable, it can not be ruled out that they are pheasants. However, the continued placement of the Grey Peacock with either Pavoninae or Perdicinae suggests it may be more likely that it is a peafowl or partridge. To resolve these questions, longer mitochondrial sequences and other markers from all of the species studied need to be included, as well as information from other species.
Acknowledgements
I would like to thank D. Bush, D. Schoening, P. Schofield, T. Matthys, D. Baker, and O. Meidinger for their donation of blood or pin feather samples.
Project Partners
Dr. C. Strobeck (University of Alberta), C. Davis (University of Alberta), & I. Delisle (University of Alberta)
References
Allen, G. A. Jr. 1966. Ruffed Pheasants: Genus Chrysolophus (Lady Amherst, Golden, Dark-Throated Golden, Yellow Golden, & Salmon Golden). Game Bird Breeders, Pheasant Fanciers and Aviculturists’ Gazette 1: 6-13.
Amador, A. G. Galliformes: Gallinaceous Birds (visited/last modified July 14, 1999) http://www.il-st-acad-sci.org/kingdom/bird002.html.
Beebe, W. 1990. A Monograph of the Pheasants: Volumes I & II. pp. 3-212. New York: Dover Publications Inc.
Beebe, W. 1990. A Monograph of the Pheasants: Volumes III & IV. pp. 3-201. Dover Publications Inc, New York.
Delacour, J. 1977. The Pheasants of the World. Spur Publications Saiga Publishing Co, Surrey, England and the World Pheasant Association, Suffolk, England.
Ghigi, A. 1966. Ghigi’s Yellow Golden Pheasant (Chrysolophus pictus luteus), and Salmon Golden Pheasant (Chrysolophus pictus infuscatus). Game Bird Breeders, Pheasant Fanciers and Aviculturists’ Gazette 1: 13-17.
Johnsgard, P. A. 1986. The Pheasants of the World. pp. 3-280. Oxford University Press, New York.
Johnsgard, P. A. 1988. The Quails, Partridges, and Francolins of the World. pp. 3-9 and 111-121. Oxford University Press, New York.
Kimball, R. T., Braun, E. L., and Ligon, J. D. 1997. Resolution of the phylogenetic position of the Congo peafowl, Afropavo congensis: a biogeographic and evolutionary enigma. Proceedings of the Royal Society of London 264: 1517-1523.
Kimball, R. T., Braun, E. L., Zwartjes, P. W., Crowe, T. M., and Ligon, J. D. 1999. A Molecular Phylogeny of the Pheasants and Partridges Suggests that these Lineages are not Monophyletic. Molecular Phylogenetics and Evolution 11(1): 38-54.
Kimball, R. T. and Ligon, J. D. 1999. Evolution of avian plumage dichromatism from a proximate perspective. The American Naturalist 154(2): 182-193.
Randi, E. 1996. A Mitochondrial Cytochrome B Phylogeny of the Alectoris Partridges. Molecular Phylogenetics and Evolution 6(2): 214-227.
Randi, E., Fusco, G., Lorensini, R., and Crowe, T. M. 1991. Phylogenetic Relationships and Rates of Alloenzyme Evolution within the Phasianidae. Biochemical Systematics and Ecology 19(3): 213-221.
Randi, E., Lucchini, V., Armijo-Prewitt, T., Kimball, R. T., Braun, E. L., and Ligon, J. D. 2000. Mitochondrial DNA Phylogeny and Speciation in the Tragopans. The Auk 117(4): 1003-1015.
Randi, E., Lucchini, V., Hennache, A., Kimball, R. T., Braun, E. L., and Ligon, J. D. 2001. Evolution of the Mitochondrial DNA Control Region and Cytochrome b Genes and the Inference of Phylogenetic Relationships in the Avian GenusLophura (Galliformes). Molecular Phylogenetics and Evolution 19 (2): 187-201.